

 Roger Iziga-Goicochea, Hozmara Rocío Torres-Acosta
Roger Iziga-Goicochea, Hozmara Rocío Torres-AcostaA principios del año 1900, los investigadores descubrieron que las moléculas solubles en lípidos ingresaban a la célula más rápidamente que las moléculas solubles en agua, lo que sugirió que los lípidos son los componentes de la membrana plasmática.
Posteriomente, análisis químicos revelaron que la membrana contenía fofolípidos.
Gorter y Grendel (1925) hayaron que la cantidad de fosfolípidos extraidos a partir de glóbulos rojos de la sangre fue suficiente como para formar una bicapa; ellos sugirieron que las colas no polares de los fosfolípidos se ubicaban hacia adentro y las cabezas polares hacia afuera.
Para explicar la permeabilidad de la membrana a las sustancias no lipídicas, Danielli y Davson (1940) propusieron el modelo del "sandwich", con una bicapa de fosfolípidos ubicada entre capas de proteína.
Robertson (1950) propuso que las proteínas se encontraban "incrustadas" en una membrana externa y que todas las membranas celulares tenian una composición similar, lo que se denominó el modelo de "unidad de membrana".
Investigaciones adicionales mostraron una gran diversidad en la estructura y función de la membrana celular.
En 1972, Singer y Nicolson introdujeron el modelo del mosaico fluido que se acepta hoy.
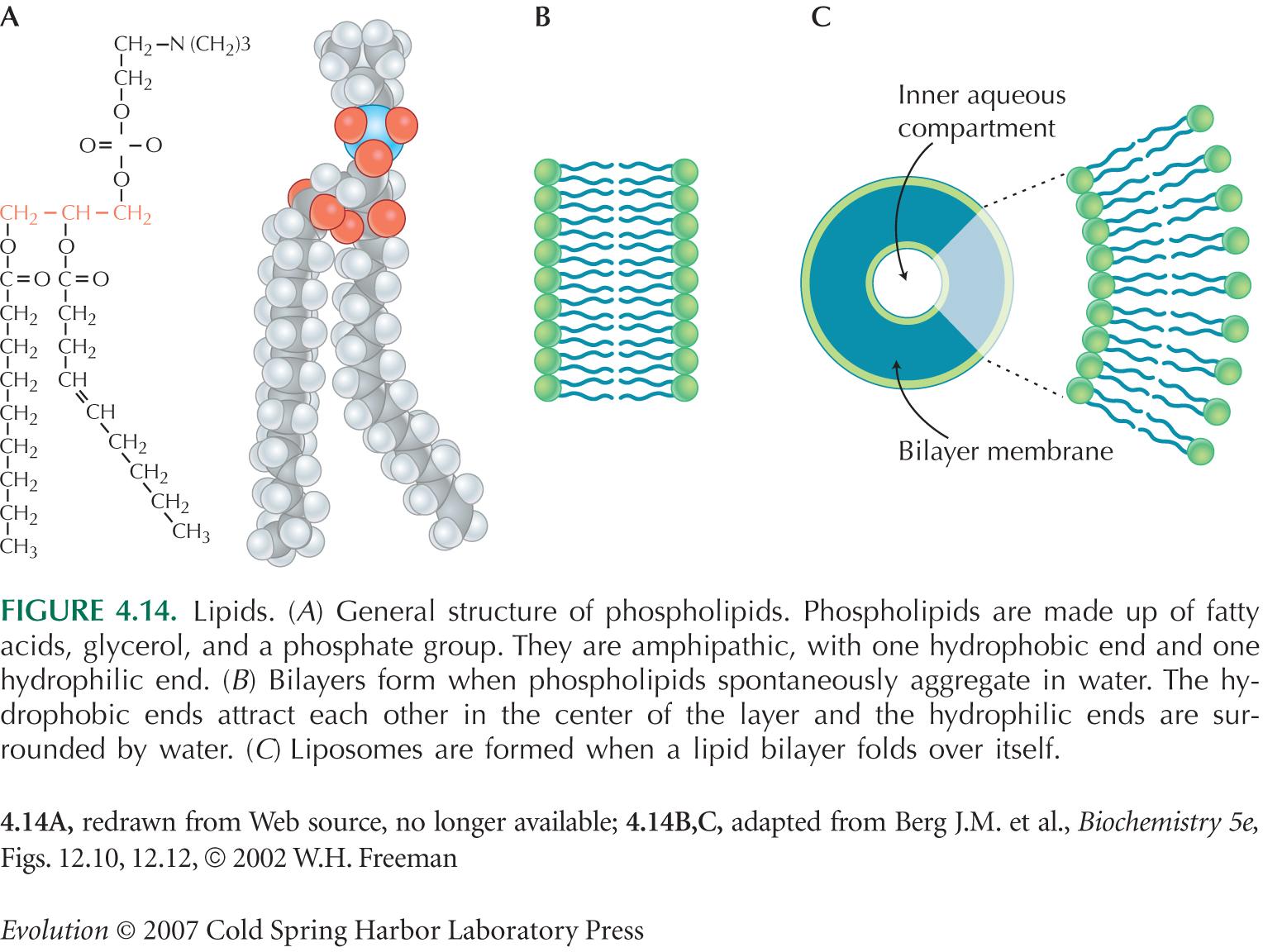
La membrana plasmática está formada por una bicapa de fosfolípidos en la que las proteínas se encuentran "incrustadas". Estas proteinas están dispersas en la membrana siguiendo un patrón irregular que varía de membrana en membrana.
La membrana plasmática es una bicapa de fosfolípidos con proteínas "incrustadas". Los fosfolípidos tienen una región hidrofílica y otra hidrofóbica; las colas no polares (hidrofóbicas) se ubican hacia el interior de la membrana, las cabezas polares (hidrofílicas) se ubican hacia el exterior en contacto con el fluido intra y extracelular. Las proteínas forman un patrón en forma de mosaico en la membrana.
El colesterol es un liupoido que se encuentra en la membrana plasmatica de los animales dando firmeza y fortaleza a la membrana.
Los glucolipidos tienen una estructura similar a la de los fosfolipidos excepto por que la cabeza hidrofilica esta formada por algun tipo de azucar; éstos cumplen funciones de proteccion, de reconocimiento entre otros.
las glicoproteinas tienen unidas cadenas de carbohidratos que se proyectan externamente.
la membrana plasmatica es asimetrica; los glicolipidos y las proteinas se encuentran ubicados hacia el exterior y los filamentos del citoesqueleto se unen a las proteinas solamente en la superficie interna de la membrana.
En las células animales, el glucocalix es una "cubierta de azúcares" formado por cadenas de carbohidratos.
Las células son únicas debido en parte a que tienen una gran variedad de cadenas de carbohidratos (la huella digital celular). El sistema inmunológico reconoce tejidos extraños que tienen cadenas inapropiadas de carbohidratos. Las cadenas de carbohidratos determinan los grupos sanguíneos A, B, AB y O en los humanos.
A la temperatura corporal, la bicapa de fosfolípidos tiene la consistencia del aceite de olivo. Mientras mayor sea la concentración de residuos de ácidos grasos insaturados, mayor será la fluidez de la bicapa.
En cada monocapa, las cadenas de carbohidratos se pueden mueven de lado a lado con pequeños movimientos y las moléculas de fosfolípidos puede desplazarse de un lado a otro en la monocapa.
La fluidez de la bicapa de fosfolípidos le permite a la célula ser flexible.
Las proteínas se mantienen en sus posiciones en la membrana por que están unidas y ancladas a los filamentos del citoesqueleto; muchas otras proteínas se pueden desplazar en la bicapa fluida.
Las proteínas de membrana determinan muchas de las funciones de la membrana. Las proteínas que forman canales de membrana le permiten a una molécula atravesar libremente la membrana (e.g., canales de paso de Cl-). Las proteínas transportadoras interactúan selectivamente con una molécula específica de tal manera que ésta pueda cruzar la membrana (e.g., la bomba Na+-K+). Las proteínas de reconocimiento celular son glucoproteínas que permiten al sistema inmunológico distinguir entre células extrañas o invasoras y las células propias del cuerpo. Las proteínas receptoras tienen una forma determinada de manera que una molécula específica pueda unirse a ellas. Las proteínas enzimáticas desempeñan funciones específicas en las reaccioes metabólicas.
Pequeñas moleculas lipídicas no cargadas eléctricamente (alcohol, oxígeno) atraviesan la membrana sin dificultad.
Pequeñas moléculas polares (dióxido de carbono, agua) se mueven en función del gradiente de concentración, desde un área de mayor concentración a una de menor concentración.
Los iones y otras moléculas cargadas eléctricamente no pueden atravesar directamente los componentes hidrofóbicos de la bicapa por lo que usualmente hacen uso de una proteína transportadora para atravesarla.
Los mecanismo de transporte pasivo y activo mueven las moléculas a través de la membrana.
El transporte pasivo permite el moviminto de las molécuals a través de la membrana sin gasto de energía; este proceso incluye la difusión y el transporte facilitado.
El transporte activo requiere de proteínas transportadoras y el uso de energía (ATP) para mover las moléculas por la membrana; este proceso incluye el transporte activo, exocitosis, endocitosis y pinocitosis.
Una solución contiene un soluto, usualmente un sólido, y un solvente , usulamente un líquido.
En el caso de la difusión de un colorante en el agua, el colorante es el soluto y el agua es el solvente.
Una vez que el soluto es distribuido uniformente en el solvente, los movimientos al azar de las moléculas siguen ocurriendo sin ningún cambio neto en las concentraciones del soluto en solución.
Las propiedades físicas y químicas de las membranas permiten solo a unos pocos tipos de moléculas atravesarla por difusión.
Los gases se difunden por la bicapa de lípidos; e.g., el movimiento de oxígeno desde los alveolos a la sangre de los capilares pulmonares depende de la concentración de oxígeno en los alveolos.
La temperatura, presión, corrientes eléctricas y el tamaño molecular influyen en la velocidad de difusión.
La ósmosis es la difusión de agua a través de una membrana selectivamente permeable.
La osmosis se puede ilustrar con el siguiente ejemplo: Una membrana semipermeable separa dos soluciones. Hacia un lado de la membrana hay más concentración de agua (menor porcentaje de soluto), mientras que en el otro lado hay menos agua (mayor porcentaje de soluto). La membrana no permite el paso del soluto; el agua entra pero el soluto no puede salir. La menbrana permite la entrada de agua con un movimiento neto desde la zona de mayor concentración hacia la zona de menor concentración. La presión osmótica aumenta en uno de los lados del sistema (en donde hay mayor porcentaje de soluto) debido a la ósmosis.
La presión osmótica produce que el agua sea abosorbida por los riñones y sea extraida de los fluidos tisulares.
La tonicidad es la fuerza de una solución respecto de la presión osmótica. Las soluciones isotónicas son aquellas en donde la concentración relativa de los solutos en dos soluciones son iguales; una solucion salina al 0.9% es utilizada en inyecciones debido a que es isotónica en relación a los globulos rojos.
Una solucion hipotónica tiene una concentración de soluto que es menor respecto de otra solución; cuando una célula es colocada en una solución hipotónica, el agua entra en la célula y ésta puede sufrir una citólisis (la célula estalla).
El hinchamiento de una célula vegetal en una solución hipotónica genera una presión de turgencia; ésto le permite a la planta mantenerse en posición erecta o erguida.
Una solución hipertónica tiene una concentración de soluto mucho mayor respecto a otra solución; cuando una célula es colocada en una solución hipertónica, se deshidrata perdiendo tamaño y forma (una condición llamada crenación).
La plasmólisis es la disminución del citoplasma debido a un proceso de ósmosis en una solución hipertónica; si la vacuola central pierde agua, la membrana plasmática se aleja de la pared celular retrayendose al interior.
La membrana celular impide el paso de muchas sustancias, pero otras entran o salen a velocidades rápidas.
Las proteínas transportadoras son proteínas de membrana que se unen y transportan un solo tipo de molécula o ion; se cree que estas proteínas sufren un cambio en su forma que les permite transportar a las moléculas a través de la membrana.
El transporte facilitado es el transporte de un soluto específico en favor o en contra de su gradiente de concentración (que puede ser alto o bajo), y es facilitado por una proteína transportadora; la glucosa y los aminoácidos se mueven a través de la membrana de esta manera.
El transporte activo es el transporte de un soluto específico a través de la membrana celular en favor o en contra de su gradiente de concentración (que puede ser alto o bajo) utilizando la energía celular en forma de ATP.
El iodo se concentra en las células de la glándula tiroides, la glucosa es completamente absorbida en el tracto digestivo, y el sodio es principalmente reabsorbido en los tubulos renales.
El transporte activo requiere de proteínas transportadoras y ATP; por ello las células deben tener un alto número de mitocondrias cerca de la membrana donde ocurre el transporte activo.
Las proteínas involucradas en el transporte activo se denominan frecuentemente "bombas"; la bomba de sodio-potasio es un importante sistema transportador en las células nerviosas y musculares.
La sal (NaCl) atraviesa la membrana celular debido a que los iones sodio son bombeados y el ion cloro es atraído hacia el ion sodio, y por simple difusión atraviesa canales específicos en la membrana.
Transporte Asistido en la Membrana
En la exocitosis, una vesícula formada en el aparato de golgi se fusiona con la membrana celular mientras ocurre la secreción de su contenido; la insulina sale de las células secretoras de insulina por este método.
Durante la endocitosis, la célula atrapa e ingresa sustancias mediante la formación de vesículas cuando la membrana celular se invagina hacia el interior como en la fagocitosis, pinocitosis o endocitosis mediada por receptores.
En la fagocitosis, las células "ingieren" grandes partículas (e.g., bacterias), formando una vesícula endocítica.
La fagocitosis la realizan comunmente células del tipo ameboide (e.g., amebas y macrofagos).
Cuando la vesícula endocítica se fusiona con un lisosoma, ocurre la digestión de las sustancias internalizadas.
La pinocitosis se produce cuando se forman vesículas alrededor de un líquido o partículas muy pequeñas; este fenomeno es visible solo con la microscopía electrónica.
La endocitosis mediada por recepetores es una forma de pinocitosis y se produce cuando macromoléculas específicas se unen a recepotres en la membrana celular.
Los receptores proteinicos cambian su forma tridimensional para adaptarse y unirse de forma específica con las sustancias (vitaminas, hormonas, lipoproteínas, etc.); se encuentran en una determinada ubicación en la membrana celular.
En el sitio de ubicación se encuentra una invaginación con una cubierta de proteínas fibrosas hacia el lado citoplasmático; cuando la vesícula no esta cubierta con sus proteínas fibrosas, se puede fusionar con un lisosoma.
Las invaginaciones en la membrana están asociadas con el intercambio de sustancias entre las células (e.g., sangre materna y fetal).
Este sistema es selectivo y más eficiente que la pinocitosis; es importante en el movimiento de sustancias desde la sangre materna hacia el feto.
El colesterol (transportado en una molécula denominada lipoproteína de baja densidad-LDL) ingresa a una célula desde el torrente sanguíneo via los receptores de membrana en invaginaciones recubiertas de proteínas ; en la hipocolesterolemia familiar, los receptores LDL no se pueden unir a las invaginaciones recubiertas de la membrana y el exceso de colesterol se acumula en el sistema circulatorio.
Modificación de las Superficies Celulares
Superficies Celulares en Animales
Las uniones entre las células son puntos de contacto que les permiten comportarse de una manera coordinada.
Las uniones de anclaje unen mecánicamente células adyacentes.
En las uniones de adhesión, las placas citoplásmicas internas, firmemente unidas al citoesqueleto dentro de cada célula dan gran resistencia y permiten que el tejido se estire (e.g., en el corazón, estómago, vejiga).
En los desmosomas, un simple punto de unión entre células adyacentes conecta los citoesqueletos de dichas células.
En las uniones estrechas (tight junctions), las proteínas de la membrana celular se unen a la manera de un cierre o ziper ajustado; estas uniones mantienen unidas a las células tan estrechamente que los tejidos se convierten en barreras (e.g., epitelio de revestimiento del estómago, túbulos renales, barrera hemato cefálica).
Una unión comunicamente (gap junction) permite a las células comunicarse; se forma cuando dos canales de membrana idénticos se unen.
Las uniones les dan solidez y fortaleza a las células involucradas, y permiten el movimiento de pequeñas moléculas e iones desde el citoplasma de una célula al citopalsma de otra célula.
Las uniones comunicantes permiten el flujo de iones en la contracción del músculo cardiaco y del músculo liso.
La matriz extracelular es semejante a una red de polisacáridos y proteínas que producen las células animales.
El colágeno le proporciona a la matriz fortaleza y elasticidad lo que le da flexibilidad.
Las proteinas fibronectina y laminina se unen a los receptores de membrana en las células y permiten la comunicación entre la matriz extracelular y el citoplasma; estas proteínas tambien forman "autopistas" que dirigen la migración de las células durante el proceso de desarrollo.
Los proteoglicanos son glicoproteínas que proveen una base de gel que une las varias proteínas en la matriz, y principalmente regulan a las proteínas de señal que se unen a los receptores de membrana.
Pared Celular
Las células vegetales están rodeadas por una pared celular porosa; ésta varía en grosor, dependiendo de la función de la célula.
Las células vegetales tienen una pared celular primaria compuesta del polímero celulosa, que forma una estructura a manera de microfibrillas.
Las fibras de celulosa forman una red cuyos espacios están llenos de moléculas no celulósicas.
Las pectinas permiten a la pared celular amoldarse, y son abundantes en la lámina media que mantienen unida a dos células adyacentes.
Los polisacáridos no celulósicos fortalecen la pared de células maduras.
La lignina añade fortaleza y es un componente común de las paredes celulares secundarias en plantas lenosas.
Los plasmodesmos o canales estrechos ubicados en las membranas celulares, atraviesan las paredes celulares de células vecinas e interconectan sus citoplasmas, permitiendo el intercambio directo de moléculas e iones entre las células.
Fuentes de Información:
Sadava D. et al. 2006. The Science of Biology. 8th Edition. W.H.Freeman
Mader S. 2007. Biology. 9th Edition. McGraw-Hill
Imagenes tomadas de:
http://superfund.pharmacy.arizona.edu/toxamb/images/f1-1-2-b.jpg
http://www.educa.aragob.es/iescarin/depart/biogeo/varios/BiologiaCurtis/Seccion%201/6-11.jpg














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}