



Los estudios detallados de la célula comenzaron a partir del año 1830; algunos de los científicos que contribuyeron a la comprensión de la estructura y función celular fueron Robert Brown, Matías Schleiden, Theodor Schwann y Rudolph Virchow.
La teoría celular señala que todos los organismos están compuestos de células, que éstas son las unidades estrcturales y funcionales de los organismos vivos, y que las células provienen de otras células preexistentes.
Tamaño Celular
Las células pueden tener tamaños que van desde el milímetro hasta el micrómetro. Las células necesitan un área superficial de membrana plasmática lo suficientemente grande como para permitir un adecuado intercambio de materiales con su entorno. La relación área/volumen requiere que las células sean pequeñas. A medida que las células son más grandes en volumen, el área superficial relativa al volumen disminuye.
El tamaño determina cuan activa metabólicamente puede llegar a ser una célula. Las células que necesitan un área superficial mucho mayor utilizan membranas modificadas como pliegues, microvellosidades, etc.
La Microscopía en la actualidad
Los microscopios electrónicos de transmisión (TEM) usan electrones que son enfocados por magnetos y que pasan a través de la muestra a observar.
El microscopio electrónico de escanéo (SEM) usa el enfoque de electrones a través de las muestras que son recubiertas con metales; los electrones secundarios liberados a partir del metal son capturadores por un detector que permite elaborar una imagen.
La amplificacion de la imagen es una función de la longitud de onda de la luz; a menor longitud de onda los electrones permiten una mayor amplificación de la imagen que con una mayor longitud de onda.
La resolución es la distancia mínima a la cual dos objetos pueden ser vistos como separados.
La microscopia de inmunoflorescencia utiliza anticuerpos fluorescentes para estudiar proteínas u otras moleculas orgánicas dentro de las células.
La microscopia confocal utiliza rayos laser para enfocar planos no profundos en las células; ésto forma una serie de secciones ópticas con las que un computador puede crear una imagen tridimensional.
La microscopia de contraste con sistema de video puede acentuar las regiones iluminadas u oscuras, y usa un computador para contrastar las regiones con colores no reales (falsos).
Los microscopios de campo-brillante, contraste de fase, interferencia diferencial, campo oscuro, son diferentes tipos de microscopios de luz.
Los procariotas se ubican dentro de dos dominios taxonómicos: Bacteria y Archaea. Los organismos en estos dos dominios son muy similares estructuralmente pero diferentes bioquímicamente.
Las Estructura Bacteriana
Las bacterias son extremadamente pequeñas; tienen un tamaño promedio de 1–1.5 μm de ancho y 2–6 μm de largo. Presentan tres formas básicas: esférica (cocos), abastanonada (bacilos) y en espiral (espirilos rígidos o espiroquetas flexibles).
La Envoltura Celular
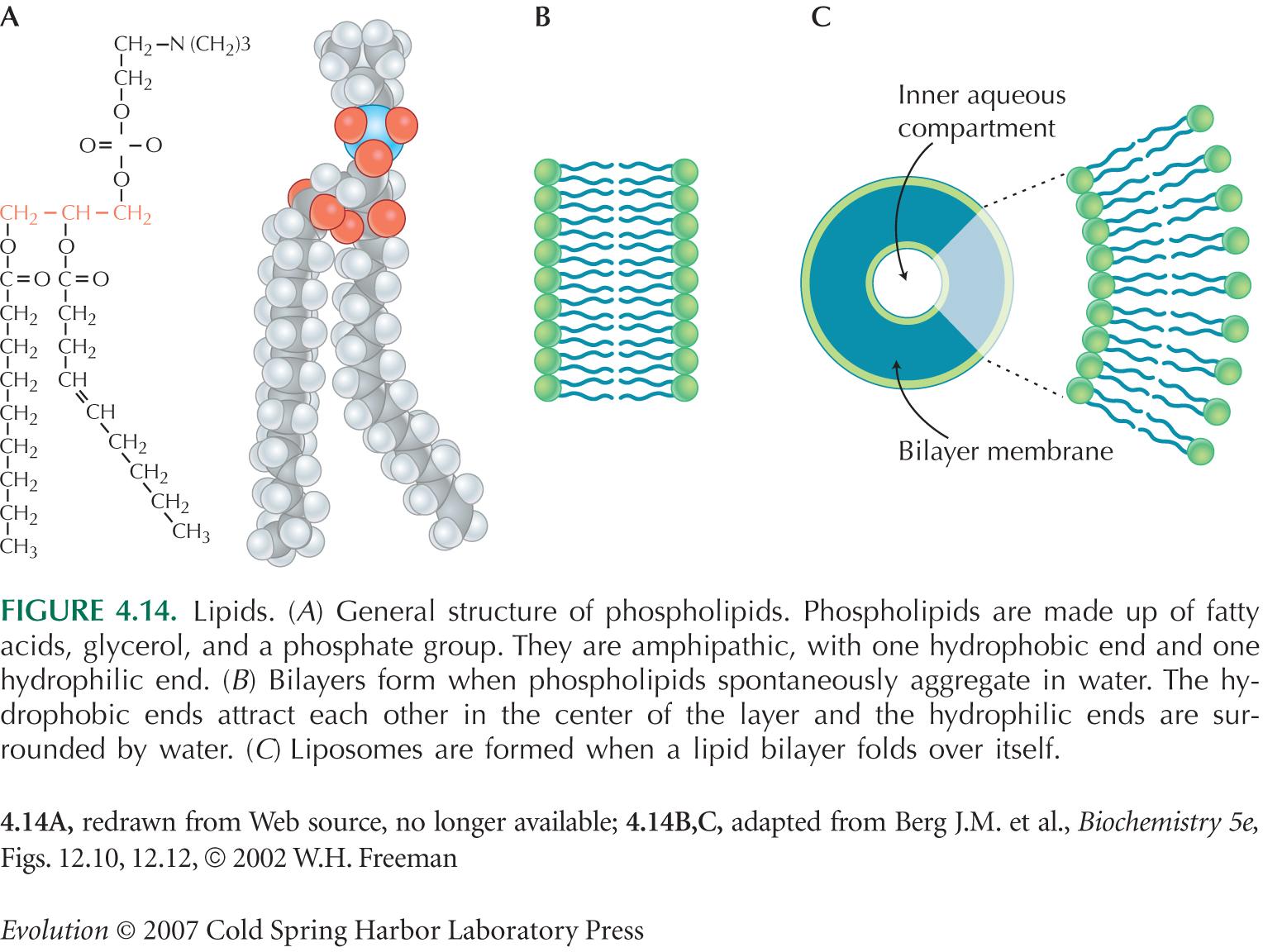
Está formada por la membrana celular o plasmática, la pared celular y el glicocalix. La membrana celular es una bicapa lipídica que contiene proteínas integrales o periféricas; ésta regula el movimiento de sustancias dentro y fuera de la célula.
La membrana celular puede formar estructuras a manera de "bolsillos o pliegues internos" denominados mesosomas, los que incrementan el área superficial interna de la membrana para su unión con las enzimas.
La pared celular mantiene la forma de la célula, y debe su fortaleza molecular a la presencia de petidogluganos.
El glicocalix es una capa de polisacáridos ubicados en la parte externa de la pared celular; se denomina cápsula si está muy bien organizada y no se remueve fácilmente, o limo si no está bien organizada y se remueve fácilmente.
Citoplasma
El citoplasma es una solución semifluida que contiene agua, moléculas orgánicas e inorgánicas y enzimas.
El nucleoide es una región que contiene una molécula de ADN circular. Los plásmidos son pequeños anillos accesorios de ADN extracromosómico.
Los ribosomas son estructuras formadas de dos subunidades de ARN y proteínas; participan en la síntesis de proteínas.
Los cuerpos de inclusión que se ubican en el citoplasma son gránulos que almacenan sustancias.
Las cianobacterias (también llamadas bacterias azul-verdosas) pueden realizar la fotosíntesis; ellas carecen de cloroplastos pero poseen tilacoides que contienen clorofila y otros pigmentos.
Las fimbrias son pequeñas estructuras semejantes a cerdas fibrosas, unidas a una superficie.
Los pelos sexuales son protruciones tubulares que salen desde una bacteria y son utilizados por éstas para traspasar el ADN de una célula a otra.
Un núcleo membranoso almacena el ADN; los núcleos de las células pueden haberse originado a partir de una invaginación de la membrana celular.
Las células eucariotas son mucho más grandes que las procariotas, y debido a ésto tienen menos área superficial por volumen.
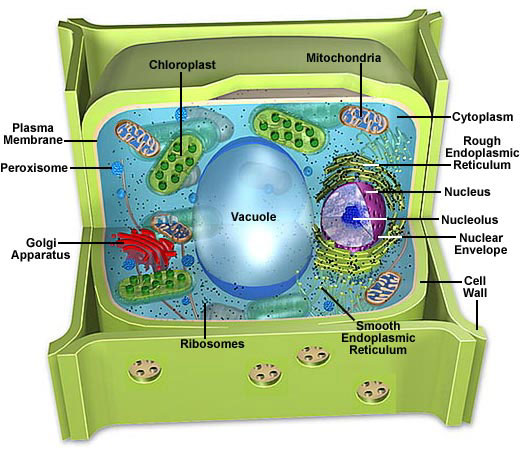
Las células eucariotas están compartimentalizadas; contienen pequeñas estructuras denominadas organelos las que realizan funciones celulares específicas.
Algunas células eucariotas (células vegetales) tienen una pared celular formada de celulosa; los plasmodesmos son canales ubicados en la pared celular que permiten la conexión del citoplasma entre células adyacentes.
Las mitocondrias (presentes en células animales y vegetales) y cloroplastos (presentes en células vegetales) están relacionados con la producción de energía. Éstos organelos no se comunican con otros organelos; ellos contienen su propio ADN y son autónomos.
El citoesqueleto está formado por una red de proteínas fibrosas que ayudan a mantener la forma y el movimiento en la célula.
Los cromosomas son estructuras semejantes a varillas que se forman durante la división celular; están compuestas de cromatina super enrollada.
El nucleolo es una "región oscura" de cromatina dentro del núcleo; es el sitio donde el ARN ribosómico (ARNr) se une con las proteínas para formar los ribosomas.
El núcleo está separado del citoplasma por una envoltura o membrana nuclear, la que contiene poros nucleares que permiten la entrada o salida de sustancias del núcleo (e.g., subunidades ribosomales, ARN mensajero, proteínas, etc.).
Los ribosomas son estructuras macromoleculares en donde se produce la síntesis de proteínas en las células. En los eucariotas, los ribosomas se pueden encontrar libres o en grupos denominados polirribosomas dentro de sus células.
Los ribosomas se unen al ARN mensajero que proviene del núcleo, el cual instruye a los ribosomas para que estos ensamblen la secuencia correcta de aminoácidos de la proteína a ser sintetizada.
El Sistema de Endomembranas
Este sistema consta de una envoltura nuclear, las membranas del retículo endoplásmico, el aparato de Golgi y diferentes tipos de vesículas.
El Retículo Endoplásmico
El retículo endoplásmico (RE) es un sistema de canales membranosos y sáculos (vesículas aplanadas) continuos con la membrana externa de la envoltura nuclear. El RE rugoso está cubierto de ribosomas hacia la cara citoplásmica; es el sitio en donde se sintetizan las proteínas que ingresan al interior del RE para ser procesadas y modificadas. El RE liso es continuo con el RE rugoso pero carece de ribosomas; alli se producen varios procesos de síntesis, detoxificación y almacenamiento; el RE liso forma vesículas de transporte.
El Aparato de Golgi
Este organelo debe su nombre a Camilo Golgi, quien lo descubrió en 1898.
El aparato de Golgi está formado por un apilamiento de sáculos ligeramente curvos. El Golgi recibe vesículas cargadas de proteínas que se desprenden desde el RE rugoso y vesículas llenas de lípidos desde el RE liso.
Las enzimas dentro del aparato de Golgi modifican los carbohidratos que fueron agregados a las proteínas en el RE (glucoproteínas); las proteínas y los lípidos son clasificadas y empaquetadas.
Las vesículas formadas a partir de la membrana de la cara mas externa del aparto de Golgi se mueven hacia diferentes lugares en la célula; en la membrana plasmática las vesículas descargan su contenido en forma de secresión, proceso denominado exocitosis debido a que las sustancias salen de la célula.
Lisosomas
Los lisosomas son vesículas rodeadas por una membrana; se originan a partir del aparato de Golgi. Los lisosomas contienen enzimas digestivas muy potentes que se encuentran en un ambiente muy ácido.
Las macromoléculas ingresan a la célula en el interior de vesículas; los lisosomaa se fusionan con las vesículas y digieren su contenido.
Los glóbulos blancos de la sangre que atrapan bacterias usan los lisosomas para digerirlas.
La autodigestión ocurre cuando los lisosomas digeren partes de la misma célula.
Los lisosomas participan en la apoptosis, o muerte celular programada, un evento normal en el desarrollo de los organismos.
Sistema de Endomembranas
Las proteínas producidas en el RE rugoso y los lípidos del RE liso son transportados en vesículas hacia el aparato de Golgi.
El aparato de Golgi modifica estos produstos (proteinas, lípidos) y luego los segrega y empaca en vesículas que van a varios destinos en la células.
Las vesículas secretorias llevan productos hacia la membrana en donde por medio de la exocitosis se produce la secresión.
Los lisosomas se fusionan con las vesículas que ingresan a la célula y digieren las macromoléculas.
Peroxisomas y Vacuolas
Los peroxisomas en las células del higado intervienen en la producción de sales biliares a partir del colesterol, así como también intervienen en la degradación de grasas.
Los peroxisomas estan presentes en las semillas en germinación donde convierten los aceites en azúcares que usan como nutrientes para el crecimiento de la planta.
La vacuola central de una celula vegetal mantienen la presión de turgencia dentro de la célula, almacena nutrientes, productos de desecho, y degrada organelos a medida que estos envejecen.
Los cloroplastos son organelos membranosos (un tipo de plastidio) en donde se produce la fotosíntesis. La fotosíntesis se puede representar mediante la siguiente ecuación:
energía solar + dióxido de carbono + agua → carbohidrato + oxígeno
Únicamente las plantas, algas y cierto tipo de bacterias, son capaces de realizar fotosíntesis.
El cloropasplasto está formado por una doble membrana organizada en sacos semejantes a discos aplanados denominados tilacoides; un conjunto de tilacoides que forman algo parecido a un apilamiento de sacos se denomina una grana.
La clorofila y otros pigmentos capturan la energía solar; las enzimas que sintetizan los carbohidrartos se ubican en los cloropalstos.
Los cloroplastos tienen ADN y ribosomas; este hecho contribuye a fortalecer la hipotesis endosimbiótica.
Otros tipos de plastidios, los que difieren en color, forma y función de los cloroplastos, incluyen a los cromoplastos y leucoplastos.
La mitocondrias esta rodeadas por una doble membrana: la membrana interna rodea la matriz y se pliega formando las denominadas crestas mitocondriales. Las mitocondrias son más pequeñas que los cloropastos y frecuentemete varian en su forma. Pueden estar fijas en una sola ubicación o pueden formar largas cadenas móviles.
Las mitcondrias contienen ribosomas y su propio ADN. La matriz de la mitocondria está llena de enzimas que degradan carbohidratos. La produccion de energía química en forma de ATP se realiza en las crestas mitocondriales.
Mas de cuarenta enfermedades diferentes que involucran a las mitocondrias han sido descritas por la ciencia.
El citoesqueleto
El citoesqueleto está constituido por una red de túbulos y filamentos interconectados; se extiende desde el núcleo hacia la membrana plasmática en los eucariotes.
La microscopia electronica muestra un citosol organizado. La microscopia de inmunoflorecencia permite identificar a las proteínas fibrosas.
Lo componentes del citoesqueleto incluyen: filamentos de actina, filamentos intermedios y microtúbulos.
Filamentos de actina
Los filamentos de actina son fibras largas y delgadas (cerca de 7 nm de diámetro) y se organizan a manera de haces o redes. Los filamentos están formados de dos cadenas de monómeros de tubulina que se retuercen entre si para formar una hélice.
Los filamentos de actina cumplen un rol estructural, formando una compleja y densa red que se ubica por debajo de la membrana plasmática; esto permite la formación de seudópodos en el movimiento ameboide.
Los filamentos de actina en las microvellosidades de las celulas intestinales permiten que las celulas se acorten o alargen en el intestino.
En las celulas vegetales, los filamentos forman caminos por los cuales los cloroplastos pueden desplazarse y circular en el interior de la célula.
Los filamentos de actina se mueven cuando interactuán con la miosina; la miosina se une a los filamentos y cuando se produce la ruptura del ATP, se induce un cambio conformacional en la molécula de miosina lo que permite empujar al filamento de actina hacia adelante.
Acciones similares se producen cuando las células se separan en el proceso de división celular.
Filamentos intermedios
Los filamentos intermedios tienen un diámetro de 8–11 nm, entre el tamaño de los filamentos de actina y los microtubulos.
Estos filamentos son estructuras semejantes a cuerdas formadas por polipéptidos fibrosos. Algunos filamentos forman parte de la envoltura nuclear; otros forman parte de la membrana plasmática y de las uniones célula a célula.
Microtúbulos
Los microtubulos estructuras a manera de pequeños cilíndros huecos (25 nm de diámetro y con una longitud de 0.2–25 μm). Están compuestos de una proteína globular denominada tubulina que tiene dos formas: tubulina α y β.
Estas dos proteínas se une y forman un dímero; los dímeros se asocian en hileras para formar las estructuras cilíndricas.
La regulación del emsamblaje de los microtúbulos está bajo el mcontrol del centro organizador de los microtúbulos (MTOC): el principal MTOC se denomina centrosoma.
Los microtubulos se organizan e irradian desde el MTOC, ayudando a mantener la forma de las células y sirviendo como caminos a través de los cuales los organelos se pueden desplazar.
A semejanza de la actina y miosina, la kinesina y dineina son motores moleculáres asociados a los microtúbulos. Diferentes tipos de kinesinas se especializan en el movimiento y transporte de algún tipo de vesícula u organelo celular.
La dineina citoplasmática es similar a la molécula de dineina que se encuentra en el flagelo.
Los Centriolos
Los centriolos son estructuras cilíndricas cortas que forma un anillo con un patrón de tripletes de microtúbulos que tienen la formula de (9 + 0).
En las celulas animales y muchos protistas, los centrosomas contienen dos centriolos que se ubican formando ángulos rectos uno respecto a otro.
Las celulas vegetales y de hongos tiene el equivalente a los centrosomas, pero no contienen centriolos.
Los centriolos forman el cuerpo basal para los cilios y flagelos.
Cílios y Flagelos
Los cílios son cortos, como numerosas proyecciones a manera de pelos que se mueven siguiendo un patrón ondulante (se encuentran en el Paramecium, recubriendo el tracto respiratorio superior en el hombre).
Los flagelos son mas largos, usualmente menos numerosos, como proyecciones que se mueven a manera de un látigo (como en el caso del espermatozoide).
Ambos tiene una estructura similar pero difieren del flagelo procariote.
Los estructuras cilíndricas rodeados de membrana encierran una matriz que contiene un cilíndro formado por 9 pares de microtúbulos que rodean a dos microtúbulos simples dando un patron de microtúbulos con la formula 9 + 2.
Los cílios y los flagelos se mueven cuando los microtúbulos se deslizan moviendose unos sobre otros. Los cilios y flagelos tienen un cuerpo basal en su base con el mismo arreglo de microtúbulos triples como los centriolos.
Los cílios y flagelos crecen por agregación de dímeros de tubulina en sus extremos o puntas.
Fuentes de Información:
Sadava D. et al. 2006. The Science of Biology. 8th Edition. W.H.Freeman
Mader S. 2007. Biology. 9th Edition. McGraw-Hill
Imagenes tomadas de:
http://micro.magnet.fsu.edu/cells/plants/images/plantcell450.jpg
http://www.ict-science-to-society.org/Pathogenomics/images/bacteria_cell.jpg



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}